摘要
背景
黑色素瘤是最致命的皮膚癌形式,由於其快速轉移而聞名。最近,有幾項研究報導了硒可以誘導黑色素瘤細胞凋亡。然而,其精確的作用機制尚待闡明。在本研究中,我們研究了硒對小鼠黑色素瘤細胞增殖、對C57BL/6小鼠腫瘤生長和轉移的影響。
方法
通過MTT試驗測量硒處理的黑色素瘤細胞增殖。通過用丙啶碘化物(PI)染色DNA分析細胞週期分佈。通過逆轉錄PCR和西方墨點法測量與細胞週期阻滯相關的mRNA和蛋白質表達。通過體內模型測量腫瘤生長和轉移。
結果
硒以劑量依賴的方式抑制了黑色素瘤細胞的增殖。硒對黑色素瘤的生長抑制與細胞週期分佈在G0/G1階段的阻滯有關。CDK2/CDK4的mRNA和蛋白質水平在硒處理下呈時間依賴性被抑制。在體內,硒並未抑制腫瘤生長;然而,在小鼠模型中,硒抑制了腫瘤轉移。
結論
這些結果表明,硒可能是一種抑制黑色素瘤細胞增殖活性的有效劑。
簡介
惡性黑色素瘤是最具侵略性的皮膚癌,由於其快速轉移和對化療和放療等抗癌治療的強大抵抗力,因此其致死率高。儘管對黑色素瘤的生理學進行了廣泛的研究,但相關的死亡率仍然很高,關鍵的治療方法仍然不足。一些報告顯示,抗氧化劑的治療對於預防黑色素瘤細胞的快速生長和轉移更為有益。特別是,抗癌藥物和抗氧化劑的結合治療比單獨治療更有效,因為結合治療通過協同作用可以降低整體毒性。
硒是一種在細胞平衡和對抗細胞應激(如活性氧物種)和DNA損傷等方面起作用的微量必需元素。Patrick等人發表了硒作為抗癌劑的功能。一些報告表明,亞硒酸鈉(許多食品補充劑中的硒來源)可以強烈誘導各種癌細胞的細胞死亡。此外,最近的研究揭示了亞硒酸鈉對黑色素瘤癌細胞具有有效的抗癌活性。儘管有關硒的抗癌和抗氧化效應的證據很多,但這種化學物質如何誘導各種癌細胞的細胞死亡的確切機制尚不清楚。
最近的研究揭示了活性氧物種在硒介導的癌細胞死亡中起著關鍵作用。活性氧物種水平的增加會在硒處理後通過誘導DNA斷裂和細胞週期抑制而增加細胞死亡。眾所周知,CDK(cyclin-dependent kinase)在與活性氧物種相關的細胞週期調節中起著關鍵作用。一些研究報告稱,硒處理誘導CDK2基因表達上調,從而激活p53,進而在小鼠哺乳動物上皮癌細胞系中誘導細胞死亡。此外,研究顯示,在用硒處理後,p53和p38通路的激活可誘導人類宮頸癌中的細胞凋亡。儘管硒對某些癌細胞的生長和轉移的影響已經被廣泛研究,但硒對黑色素瘤細胞生長和細胞遷移的抗癌效應尚不完全了解。在本研究中,我們評估了硒對小鼠黑色素瘤細胞株的抗癌效應,以及硒對B16小鼠黑色素瘤細胞的實驗性轉移模型中的抗轉移能力。這些發現為抗氧化劑治療轉移性黑色素瘤癌提供了有用的信息。
材料與方法
細胞培養和試劑 B16F10小鼠黑色素瘤細胞系在RPMI 1640培養基中培養,該培養基添加了2 mM L-谷氨酰胺,100單位/毫升青黴素,100微克/毫升鏈黴素和10%熱灭活胎牛血清。細胞在37℃下在含有5% CO2的濕潤環境中培養。細胞在生長的對數階段進行實驗。亞硒酸鈉(硒)購自Sigma(美國密蘇里州聖路易斯)。
活細胞測定
使用3-(4,5-二甲基噻唑-2-基)-2,5-二苯基四唑溴化物(MTT; Sigma, 美國密蘇里州聖路易斯)試驗測定細胞生長的抑制作用。B16F10小鼠黑色素瘤細胞種植在96孔板上(每孔種植1×10 3 個細胞),並在加入20微升MTT溶液(0.5毫克/毫升)前用0、1、2、10、25、50或100微米的硒處理72小時。在加入染料後的4小時內,使用ELISA讀取器(Bio-Tek instruments, Inc., 美國佛蒙特州溫諾斯基)在570 nm波長下測量MTT吸光度。每個實驗重複進行三次。
檢測硒誘導的細胞凋亡
為了研究硒的誘導細胞凋亡效應,收集細胞,用PBS洗涤兩次,然後懸浮於100微升Annexin V結合緩衝液(10 mM HEPES/NaOH pH 7.4,140 mM NaCl,2.5 mM CaCl2)中。添加2微升FITC標記的Annexin V(BD Pharmingen, 美國加利福尼亞州聖地亞哥)和2微升丙啶碘化物(BD Pharmingen),在室溫下在黑暗中輕輕搖晃15分鐘。最後,向每個試管中加入400微升Annexin V結合緩衝液,使用FACSCalibur(BD Pharmingen)分析細胞。
細胞週期分析
使用丙啶碘化物(PI)染色法進行細胞週期分析。在亞硒酸鈉處理後收集細胞,用PBS(2% FBS)洗涤兩次,然後在4℃冷水乙醇中固定過夜。將細胞在室溫下在PI染色液中懸浮(10微克/毫升RNase A和10微克/毫升PI在PBS中),室溫下靜置30分鐘,使用FACSCalibur(BD Pharmingen)測定DNA含量。
反轉錄聚合酶鏈反應
為了評估亞硒酸鈉對細胞週期調控基因表達的影響,亞硒酸鈉處理後收集細胞,用冷PBS洗涤兩次。使用RNeasy mini kit(Quiagen,美國加利福尼亞州瓦倫西亞)按照制造商的說明提取RNA,使用RT premix(Bioneer,南韓大田)轉錄cDNA。使用以下引物進行特定基因轉錄物的RT-PCR分析:p53(上游引物:5′-ATG ACT GCC ATG GAG GAG T,下游引物:5′-CTC GGG TGG CTC ATA AGG TA,664 bp產物),p21Waf1(上游引物:5′-CTT TGA CTT CGT CAC GGA GAC,下游引物:5′-AGG CAG CGT GAA GGT GTT TGG GG,480 bp產物),CDK2(上游引物:5′-CAC AGC CGT GGA TAT CTG GAG,下游引物:5′-TTG CGA TAA CAG GCT CCG TC,253 bp產物),CDK4(上游引物:5′-ACG CCT GTG GTG GTT ACG CT,下游引物:5′-CCA TCT CTG GCA CCA CTG AC,280 bp產物)。作為內部對照,使用β-actin(上游引物:5′-AAG AGC TAT GAG CTG CCT GA,下游引物:5′-CAG GAG GAG CAA TGA TCT TG,300 bp產物)。使用AccuPower PCR premix(Bioneer)進行PCR(24個循環,94℃下30秒,60℃下30秒,72℃下60秒)。
免疫印跡
細胞(1×10 7細胞/樣品)被刮入冷 PBS 中,離心並在 RIPA 緩衝液(EIPiS,大田,韓國)冰上裂解 30 分鐘。
每個細胞裂解物在15% SDS-PAGE條件下在還原條件下進行分離,使用半幹法將其轉移到硝酸纖維膜上,以80mA的電流進行2小時。將膜用PBS中的5%脫脂奶粉阻斷1小時,然後在1:1,000的最終稀釋度下與一抗進行2小時的孵育。在PBS-T(PBS中的0.05% Tween-20)中洗滌三次,然後在稀釋緩沖液(PBS中的5%脫脂奶粉)中與過氧化物酶標記的二抗(最終稀釋度為1:3,000)孵育1小時,然後按上述方法進行洗滌。使用增強型化學發光試劑(ECL-kit;Amersham Life Science,Braunschweig,德國)和Multiple Gel-DOC系統(Fujifilm,日本東京)進行檢測。使用以下一抗:p21 Waf1/Cip1、CDK4、CDK2和 β-肌動蛋白(來自Cell Signaling,美國麻省貝弗利)。
動物和實驗性轉移模型
6至8週齡的C57BL/6小鼠由Orient(南韓首爾)獲得。動物實驗的審查委員會審查了所有動物研究。6至8週齡的C57BL/6小鼠通過靜脈內注射0.1毫升PBS,含有0.1百萬個B16F10細胞。小鼠被隨機分為5組(每組5隻小鼠)。小鼠從第1天開始通過腹腔注射給予10微米的硒處理。第4天和第8天,小鼠以200微升PBS的形式注射硒。在第14天,殺死小鼠,計數肺表面上的黑色腫瘤結節。將肺切片固定,用血紅素-嗎啡藍染色,並在相差干涉顯微鏡下觀察。
統計分析 結果至少代表三次獨立實驗。使用學生t檢驗計算統計學意義。p值小於0.05被認為是顯著的。
結果
硒對B16F10細胞生長和細胞形態的影響
為了評估硒是否直接誘導B16F10黑色素瘤細胞的細胞死亡,將細胞處理0、1、2、10、25、50或100微米的硒。使用MTT試驗來確定硒誘導的細胞生長抑制作用。圖1A顯示,暴露於50微米的硒中的細胞在72小時內的生長顯著降低,僅為對照組的35%。我們還發現硒的生長抑制作用呈劑量依賴性。為了確認我們的結果,我們觀察了暴露於40微米硒下的B16F10細胞在24小時和48小時的形態變化。如圖1B所示,以40微米的硒處理明顯地按時間依賴性降低了B16F10黑色素瘤細胞的數量。
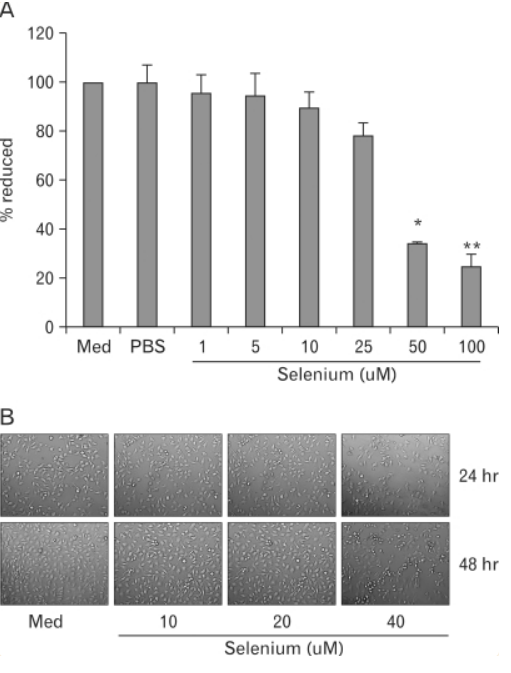
圖1. 硒對B16F10小鼠黑色素瘤細胞的細胞存活率和形態變化的影響。(A)硒抑制B16F10小鼠黑色素瘤細胞的細胞存活率(MTT試驗)。將B16F10黑色素瘤細胞(1×103)種植在96孔板上,並以不同劑量(0.1、5、10、25、50和100微米)的硒處理72小時。4小時後加入MTT溶液(100微克/孔)。 (B)將B16F10細胞(2×105)過夜種植在6孔板上進行附著,然後以不同劑量(0、10、20和40微米)的硒處理。48小時後,使用倒置顯微鏡拍攝細胞的照片。
硒對B16F10細胞的死亡和細胞週期的影響
接下來,我們通過流式細胞儀分析測量了B16F10細胞的凋亡情況。細胞培養在不同條件下處理72小時。圖2A顯示,與低劑量處理組相比,40微米硒處理後的細胞凋亡數量顯著增加。
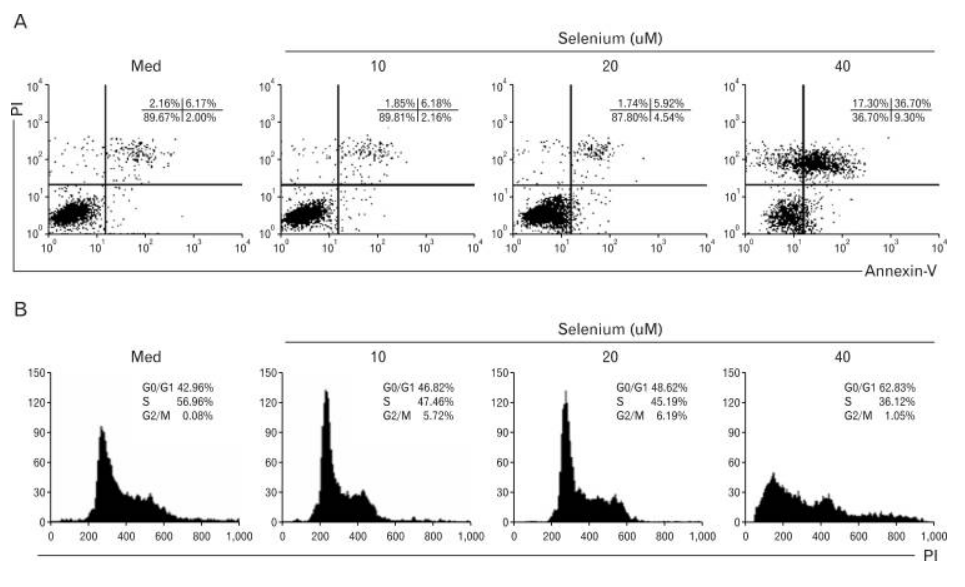
圖2. 硒誘導B16F10小鼠黑色素瘤細胞的細胞死亡和細胞週期阻滯。(A)B16F10黑色素瘤細胞(2×105)過夜種植在6孔板上進行附著。細胞暴露於不同劑量的硒(0、10、20和40微米)後孵育48小時。孵育後,使用細胞分離溶液收集細胞,並用PBS洗涤一次。使用Annexin-V-FITC/PI染色評估硒對B16F10黑色素瘤細胞的細胞毒性作用。通過流式細胞儀進行細胞死亡分析。數字表示存活細胞(Annexin-V-;PI-)細胞群體(左下象限)、凋亡(Annexin-V+)細胞群體(右下象限)以及上右象限(Annexin-V+;PI+)和上左象限(Annexin-V-;PI+)的壞死細胞群體的百分比。(B)B16F10黑色素瘤細胞經硒(0、10、20和40微米)處理後孵育48小時。使用細胞分離溶液收集細胞,用冷PBS洗涤兩次,然後離心。沉澱物在1毫升冷PBS和4毫升冷乙醇中在4℃下孵育30分鐘。然後,使用丙啶碘化物染色30分鐘,並通過流式細胞儀進行分析。圖中顯示G0/G1、S和G2/M期細胞的百分比。
為了瞭解硒誘導的細胞凋亡機制,我們使用PI染色法分析了硒處理的B16F10細胞的細胞週期。與低劑量處理組相比,添加硒(40微米)顯著增加了B16F10細胞的G0/G1阻滯(圖2B)。綜上所述,這些結果表明G0/G1細胞週期阻滯在硒誘導的細胞凋亡中起主導作用。
硒對B16F10黑色素瘤細胞中細胞週期調節分子的表達影響 為了研究硒誘導的B16F10細胞G0/G1阻滯的機制,我們調查了在硒處理後各種細胞週期調節子(如p53、p21Waf1、CDK2和CDK4)的mRNA和蛋白質表達模式。如圖3A所示,在時間依賴性的方式下,硒處理顯著降低了p53、CDK2和CDK4的mRNA表達。然而,p21Waf1的mRNA在時間依賴性的方式下增加(圖3A)。此外,在將細胞暴露於40微米的硒後,p21WAF1的蛋白質水平顯著增加,而CDK2和CDK4的蛋白質水平在時間依賴性的方式下降低(圖3B)。這個結果暗示硒誘導的細胞週期阻滯是通過一個獨立於p53的p21Waf1途徑調節的。
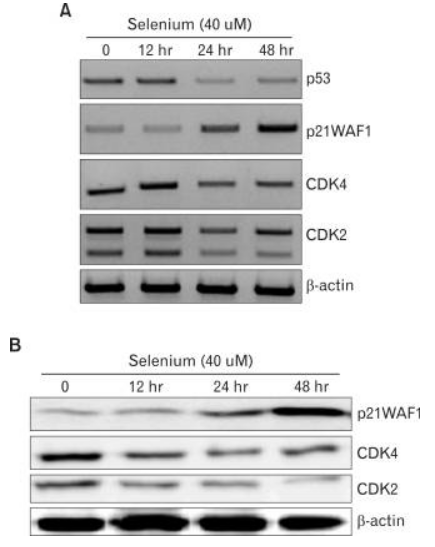
圖3. 硒對B16F10黑色素瘤細胞中細胞週期阻滯相關基因表達的影響。(A)在與硒(40微米)孵育48小時後的指定時間收集B16F10黑色素瘤細胞。從48小時處理後的細胞中提取總RNA,並按照材料和方法部分的方法製備cDNA。(B)在與硒(40微米)孵育後的指定時間收集B16F10黑色素瘤細胞。然後對細胞進行裂解,上清液進行Western blot分析,並使用抗p21Waf1/Cip1、抗CDK4和抗CDK2抗體進行免疫印跡。
硒對腫瘤生成和B16F10轉移至小鼠肺的影響
硒的治療調節了獨立於p53的途徑和細胞生長,這兩者都與腫瘤細胞生長相關。為了評估硒對黑色素瘤細胞生長的影響,我們在C57BL/6小鼠皮下接種B16F10細胞並進行硒治療後,測定了黑色素瘤細胞在體內的生長。當硒被給予同基因小鼠時,沒有觀察到腫瘤生長的嚴重退縮(圖4)。然而,接受硒治療的B16F10黑色素瘤小鼠擁有明顯減少的腫瘤負擔,通過計數肺部結節的數量來衡量(圖5A)。此外,我們使用組織化學H&E染色測試了B16F10黑色素瘤細胞轉移至帶有硒治療的小鼠肺部的情況。如圖5B和C所示,硒治療顯著減少了B16F10黑色素瘤細胞在肺部轉移斑點的數量。這些結果表明,硒在携帶黑色素瘤細胞的小鼠中促進了抗癌活性和抗轉移活性。
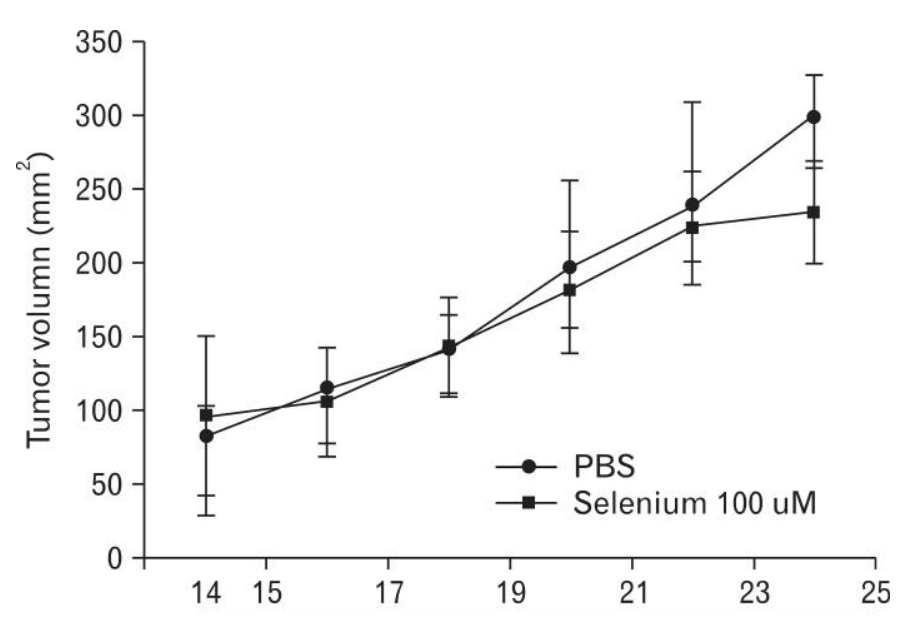
圖4. 硒對腫瘤生長的影響。B16F10黑色素瘤細胞被皮下注射到小鼠體內。硒被注射到腫瘤部位。在4天的期間內,硒被注射2次。腫瘤大小每隔一天使用游標卡尺測量一次。
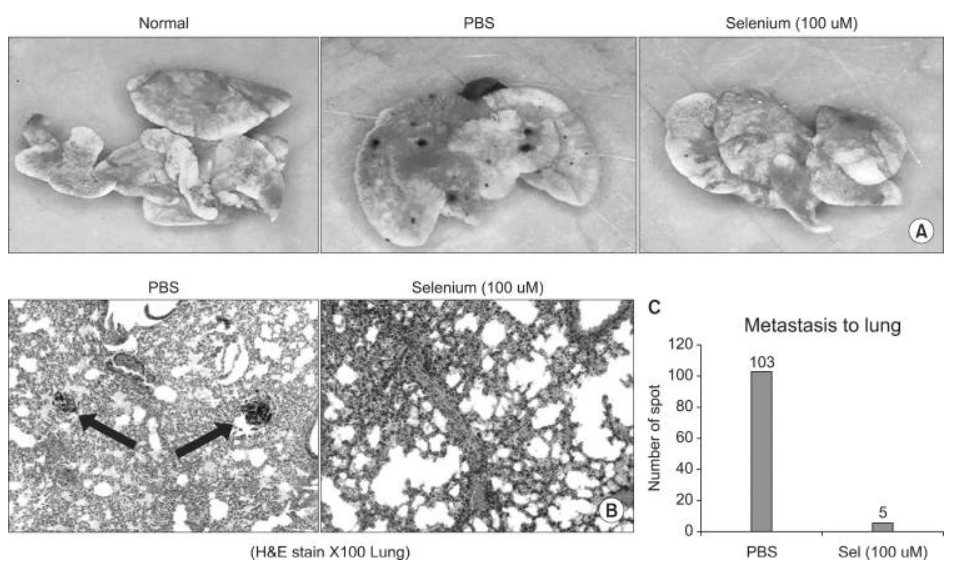
圖5. 硒對轉移至肺部的影響。腹腔注射100微米硒對B16F10黑色素瘤到肺部的實驗性轉移的影響(A)肺部的代表性照片。細胞通過眼底靜脈注射。14天後,小鼠被處理並計算B16F10黑色素瘤的肺部結節數量。(B)肺部的代表性血紅素和嗜酸性染色切片照片。箭頭指向肺部轉移瘤。組織學切片中結節佔據的肺部面積通過顯微鏡測量。(C)肺部轉移瘤的數量。
討論
切除黑色素瘤最重要的問題之一是其強大的轉移性腫瘤生長。在本研究中,我們評估了硒對控制B16F10黑色素瘤增殖和轉移的影響。眾所周知,抗氧化化合物,如抗壞血酸和硒,具有抑制癌症生長和轉移的能力(2,3)。已有報導指出,硒化合物通過添加到體外細胞培養或作為動物飲食補充劑,可以明顯減少細胞增殖和癌症發生率(6)。然而,其抗癌和抗轉移效應的詳細機制尚不清楚。
我們在體內評估了硒對B16F10黑色素瘤細胞的肺轉移的影響。有趣的是,我們的結果顯示,接受硒治療的組別在B16F10黑色素瘤細胞的轉移方面有所減少,儘管硒治療並未減少B16F10黑色素瘤細胞的腫瘤生成(圖4、5)。為了瞭解硒對B16F10黑色素瘤細胞的腫瘤生成和轉移之間的關係,我們對接受處理的細胞進行了細胞形態和凋亡的變化分析(圖1、2)。硒治療導致B16F10黑色素瘤細胞的形態變化,並以劑量依賴的方式增加了凋亡事件。這些結果表明,硒的明顯抗轉移活性可能是由於硒的凋亡活性引起的。
更有趣的是,我們的結果揭示了B16F10黑色素瘤細胞在接受硒治療後通過G0/G1細胞週期阻滯而引起細胞凋亡。這一點很重要,因為眾所周知,調節癌細胞週期是鑒定新的抗癌策略的治療目標之一。尤其重要的是G0/G1檢查點的細胞週期紊亂,這導致腫瘤生長的失調(17-21)。基於這些證據,我們關注硒誘導的凋亡中G0/G1細胞週期阻滯的機制。許多研究報導指出,p53參與了G0/G1細胞週期阻滯(22-25)。然而,Deeds等最近觀察到G0/G1阻滯是以p53獨立的方式引起的(26)。如圖3所示,我們的結果支持這些發現,即硒通過p53獨立的p21Waf1途徑誘導G0/G1阻滯。幾項研究揭示了p21WAF1在G0/G1阻滯機制中的關鍵作用,該機制涉及CDK-蛋白質複合物的結合以抑制CDK激酶活性(27)。我們發現,對黑色素瘤進行硒處理會誘導p21WAF1的表達,進而下調mRNA和蛋白質水平的CDK4和CDK2表達(圖3)。我們的研究結果表明,硒通過p53獨立的p21WAF1途徑誘導G0/G1阻滯,從而引起細胞凋亡。
總之,我們建議硒治療增強了抗轉移的能力,通過誘導p53獨立的G0/G1細胞週期阻滯促進了B16F10黑色素瘤細胞的凋亡。進一步瞭解硒治療可能為改善癌症患者的惡性程度提供有用的治療策略。
本文翻譯自論文
Hyunkeun Song,1 Indo Hur,1 Hyun-jin Park,1 Joohyung Nam,1 Ga Bin Park,1 Kyoung Hye Kong,1 Young Mi Hwang,1 Yeong Seok Kim,1 Dae Ho Cho,2 Wang Jae Lee,3 and Dae Young Hur1
“Selenium Inhibits Metastasis of Murine Melanoma Cells through the Induction of Cell Cycle Arrest and Cell Death”
Immune Netw. 2009 Dec; 9(6): 236–242. doi: 10.4110/in.2009.9.6.236
參考文獻
1. Mancianti ML, Herlyn M. Tumor progression in melanoma: the biology of epidermal melanocytes in vitro. Carcinog Compr Surv. 1989;11:369–386. [PubMed] [Google Scholar]
2. Nagao N, Nakayama T, Etoh T, Saiki I, Miwa N. Tumor invasion is inhibited by phosphorylated ascorbate via enrichment of intracellular vitamin C and decreasing of oxidative stress. J Cancer Res Clin Oncol. 2000;126:511–518. [PubMed] [Google Scholar]
3. Roomi MW, Roomi N, Ivanov V, Kalinovsky T, Niedzwiecki A, Rath M. Inhibition of pulmonary metastasis of melanoma b16fo cells in C57BL/6 mice by a nutrient mixture consisting of ascorbic Acid, lysine, proline, arginine, and green tea extract. Exp Lung Res. 2006;32:517–530. [PubMed] [Google Scholar]
4. Dua P, Ingle A, Gude RP. Suramin augments the antitumor and antimetastatic activity of pentoxifylline in B16F10 melanoma. Int J Cancer. 2007;121:1600–1608. [PubMed] [Google Scholar]
5. Combs GF, Jr, Gray WP. Chemopreventive agents: selenium. Pharmacol Ther. 1998;79:179–192. [PubMed] [Google Scholar]
6. Patrick L. Selenium biochemistry and cancer: a review of the literature. Altern Med Rev. 2004;9:239–258. [PubMed] [Google Scholar]
7. Han B, Wei W, Hua F, Cao T, Dong H, Yang T, Yang Y, Pan H, Xu C. Requirement for ERK activity in selenium-induced apoptosis of acute promyelocytic leukemia-derived NB4 cells. J Biochem Mol Biol. 2007;40:196–204. [PubMed] [Google Scholar]
8. Kim EH, Sohn S, Kwon HJ, Kim SU, Kim MJ, Lee SJ, Choi KS. Selenium induces superoxide-mediated mitochondrial damage and subsequent autophagic cell death in malignant glioma cells. Cancer Res. 2007;67:6314–6324. [PubMed] [Google Scholar]
9. Wang H, Yang X, Zhang Z, Xu H. Both calcium and ROS as common signals mediate Na(2)SeO(3)-induced apoptosis in SW480 human colonic carcinoma cells. J Inorg Biochem. 2003;97:221–230. [PubMed] [Google Scholar]
10. Kramer GF, Ames BN. Mechanisms of mutagenicity and toxicity of selenium (Na2SeO3) in salmonella typhimurium. Mutat Res. 1988;201:169–180. [PubMed] [Google Scholar]
11. Kim T, Jung U, Cho DY, CHung AS. Se-methylselencysteine induces apoptosis through caspase activation in HL-60 cells. carcinogenesis. 2001;22:559–565. [PubMed] [Google Scholar]
12. Jung U, Zheng X, Yoon SO, Chung AS. Se-methylselenocysteine induces apoptosis mediated by reactive oxygen species in HL-60 cells. Free Radic Biol Med. 2001;31:479–489. [PubMed] [Google Scholar]
13. Sherr CJ. cancer cell cycles revisited. Cancer Res. 2000;60:3689–3695. [PubMed] [Google Scholar]
14. Kaeck M, Lu J, Strange R, Ip C, Ganther HE, Thompson HJ. Differential induction of growth arrest inducible genes by selenium compounds. BioChemical Pharmacology. 1997;53:921–926. [PubMed] [Google Scholar]
15. Rudolf E, Rudolf K, Cervinka M. Selenium activates p53 and p38 pathways and induces caspase-independent cell death in cervical cancer cells. Cell biol Toxicol. 2008;24:123–241. [PubMed] [Google Scholar]
16. Vermeulen K, Van Bockstaele DR, Berneman ZN. The cell cycle: a review of regulation, deregulation and therapeutic targets in cancer. Cell Prolif. 2003;36:131–149. [PMC free article] [PubMed] [Google Scholar]
17. Albino AP, Juan G, Traganos F, Reinhart L, Connolly J, Rose DP, Darzynkiewicz Z. Cell cycle arrest and apoptosis of melanoma cells by docosahexaenoic acid: association with decreased pRb phosphorylation. Cancer Res. 2000;60:4139–4145. [PubMed] [Google Scholar]
18. Tsai J, Lee JT, Wang W, Zhang J, Cho H, Mamo S, Bremer R, Gillette S, Kong J, Haass NK, Sproesser K, Li L, Smalley KS, Fong D, Zhu YL, Marimuthu A, Nguyen H, Lam B, Liu J, Cheung I, Rice J, Suzuki Y, Luu C, Settachatgul C, Shellooe R, Cantwell J, Kim SH, Schlessinger J, Zhang KY, West BL, Powell B, Habets G, Zhang C, Ibrahim PN, Hirth P, Artis DR, Herlyn M, Bollag G. Discovery of a selective inhibitor of oncogenic B-Raf kinase with potent antimelanoma activity. PNAS. 2008;105:3041–3046. [PMC free article] [PubMed] [Google Scholar]
19. Wang CC, Chiang YM, Sung SC, Hsu YL, Chang JK, Kuo PL. Plumbagin induces cell cycle arrest and apoptosis through reactive oxygen species/c-Jun N-terminal kinase pathways in human melanoma A375.S2 cells. Cancer Letters. 2008;259:82–98. [PubMed] [Google Scholar]
20. An WW, Wang MW, Tashiro S, Onodera S, Ikejima T. Mitogen-activated protein kinase-dependent apoptosis in norcan-tharidin-treated A375-S2 cells is proceeded by the activation of protein kinase C. Chin Med J (Engl) 2005;118:198–203. [PubMed] [Google Scholar]
21. Mhaidat NM, Zhang XD, Allen J, Avery-Kiejda KA, Scott RJ, Hersey P. Temozolomide induces senescence but not apoptosis in human melanoma cells. Br J Cancer. 2007;97:1225–1233. [PMC free article] [PubMed] [Google Scholar]
22. Kagawa S, Fujiwara T, Hizuta A, Yasuda T, Zhang WW, Roth JA, Tanaka N. p53 expression overcomes p21WAF1/CIP1-mediated G1 arrest and induces apoptosis in human cancer cells. Oncogene. 1997;15:1903–1909. [PubMed] [Google Scholar]
23. Jiang C, Hu H, Malewiez B, Wang Z, Lü J. Selenite-induced p53 Ser-15 phosphorylation and caspase-mediated apoptosis in LNCaP human prostate cancer cells. Mol Cancer Ther. 2004;3:877–884. [PubMed] [Google Scholar]
24. Traynor NJ, McKenzie RC, Beckett GJ, Gibbs NK. Selenomethionine inhibits ultraviolet radiation-induced p53 transactivation. Photodermatol Photoimmunol Photomed. 2006;22:297–303. [PubMed] [Google Scholar]
25. Hahm E, Jin DH, Kang JS, Kim YI, Hong SW, Lee SK, Kim HN, Jung da J, Kim JE, Shin DH, Hwang YI, Kim YS, Hur DY, Yang Y, Cho D, Lee MS, Lee WJ. The Molecular Mechanisms of Vitamin C on Cell Cycle Regulation in B16F10 Murine Melanoma. J Cell Biochem. 2007;102:1002–1010. [PubMed] [Google Scholar]
26. Deeds L, Teodorescu S, Chu M, Yu Q, Chen C. A p53-independent G1 cell cycle checkpoint induced by the suppression of protein kinase C alpha and theta isoforms. J Biol Chem. 2003;278:39782–39793. [PubMed] [Google Scholar]
27. Peter M, Herskowitz I. Joining the complex: Cyclindependent kinase inhibitory proteins and the cell cycle. Cell. 1994;9:181–184. [PubMed] [Google Scholar]
